ANTECEDENTES
El aguacate exhibe un crecimiento rítmico, con dos o más brotaciones por año, alternando con períodos cortos de descanso. Los flujos de crecimiento pueden ser vegetativos o reproductivos. La brotación vegetativa permite al árbol aumentar o renovar su capacidad fotosintética y también generar yemas nuevas que, posteriormente, originan flujos reproductivos.. Los flujos reproductivos son aquellos brotes o crecimientos que dan lugar a la floración, pueden ser indeterminados o determinados (Alcaraz et al., 2013). Los indeterminados están constituidos por la inflorescencia y sus correspondientes hojas, mientras que los determinados carecen de hojas.

A. Inflorescencias indeterminadas; B. Inflorescencias determinadas de P. americana cv. Hass. Fuente: Alcaraz et al. (2013).
El aguacate crece principalmente en tres zonas climáticas: climas fríos, semiáridos, con lluvias predominantes en invierno (California, Chile, Israel); climas subtropicales húmedos con lluvias predominantes en verano (este de Australia, México y Sudáfrica), y climas tropicales y semitropicales con lluvias predominantes en verano (Brasil, Florida, Indonesia). Está dividido, además, en tres razas ecológicas: Mexicana, Guatemalteca y Antillana. Los cultivares dentro de cada raza presentan, generalmente, respuestas similares a las condiciones climáticas y también edáficas. Sin embargo, hay diferencias entre las razas y entre cultivares respecto a su adaptabilidad a las condiciones ambientales (Whiley y Shaffer, 1994), como es el caso del cv. ´Hass`, híbrido entre la raza Mexicana y Guatemalteca, que presenta características intermedias entre ambas.
A grandes rasgos, las condiciones ideales de temperatura para esta especie están en torno a los 25-30ºC para las diurnas, y entre 15 y 20ºC para las nocturnas. Las temperaturas por encima de los 36ºC causan serios daños, particularmente en la fecundación y el cuajado, siendo importante que ocurra un período de frío (alrededor de 10ºC) en invierno para estimular la inducción floral (Galán, 1990).
La actividad fotosintética es un indicador del crecimiento y la productividad de un cultivo. El crecimiento y la producción dependen marcadamente del reparto de carbohidratos. Aumentar la producción en especies subtropicales de fructificación poliaxial terminal, como es el caso del aguacate, plantea un desafío para el manejo agronómico, ya que el árbol presenta una tendencia natural al crecimiento vegetativo que resulta en una mayor asignación de materia seca a éste en detrimento del desarrollo de órganos reproductivos (Wolstenholme, 1990). Factores medioambientales tales como la luz, temperatura y concentración de CO2, afectan la fotosíntesis, la respiración y el reparto de carbohidratos.
El reparto de fotoasimilados está regulado por las interacciones fuente-sumidero. Las fuentes son exportadores y los sumideros importadores netos de fotoasimilados (Ho, 1988). El orden de prioridad de la demanda es función de la tasa de crecimiento (actividad del sumidero) y del tamaño del sumidero (número de frutos). El orden, generalmente, es: semilla > pulpa de los frutos = ápices de brotes y hojas > cambium > raíces > tejidos de almacenamiento (Wolstenholme, 1990). Las hojas jóvenes, mientras se hallan en expansión, son fuertes sumideros que compiten con otros órganos demandantes de la planta hasta que alcanzan su tamaño definitivo, momento en que se convierten en exportadoras netas (Ho, 1988).
La disponibilidad de luz incidente es el factor que, probablemente, ejerce la mayor influencia sobre la fotosíntesis en un huerto frutal. En el aguacate, hacia el final del crecimiento de brotes en primavera, la transmisión de luz hacia la zona de fructificación se reduce a un 40% respecto de la plena iluminación, y a distancias de 0,5 y 1,0 m dentro de la copa desde la zona de fructificación, ésta se reduce a 14% y 10%, respectivamente. Hacia finales del crecimiento de brotes de verano, la transmisión de la luz a la zona de fructificación con plena iluminación ha disminuido a un 13%, y en los puntos internos (0,5 y 1,0 m) a 9,7% y 6,3%, respectivamente (Whiley et al., 1992).
La intensidad y duración de la iluminación son factores determinantes de la floración (Coutanceau, 1964), y es de amplio conocimiento que la floración y fructificación son menos abundantes a la sombra que bajo plena luz (Meyer, 1960). Cuando la iluminación es baja, respecto de sus requerimientos, el crecimiento vegetativo se reduce, tanto en número como en longitud de los brotes, así como en el tamaño de las hojas, resultando en un menor desarrollo del árbol y una menor actividad fotosintética. De este modo, en el interior del árbol se originan numerosas ramificaciones y la densidad de ramillas exteriores reduce la iluminación, por tanto, la floración en su interior; sólo la parte exterior de la copa con iluminación adecuada presenta floración satisfactoria (Coutanceau, 1964).
FENOLOGÍA Y DESARROLLO
El aguacate se caracteriza por tener un crecimiento rítmico monopodial, es decir, con un crecimiento de una yema vegetativa terminal del eje central de cada brote que permanece y continúa su desarrollo año tras año, y es un ejemplo del modelo arquitectónico de Rauh, uno de los más frecuentes de las zonas templadas y tropicales.
El tronco forma ramas que son morfogenéticamente idénticas al tronco y las flores se originan lateralmente sin tener un efecto sobre el crecimiento de los brotes, aunque en algunos brotes existen flores en posición terminal, siendo el crecimiento subsecuentemente simpodial (Thorp y Sedgley, 1992).
Las yemas pueden ser axilares o apicales. El árbol crece principalmente desde las yemas apicales, debido a que las yemas axilares se desprenden o permanecen en estado latente (Calabrese, 1992). El vigor del crecimiento completo del árbol y la producción de fruta dependen del tiempo y extensión de los eventos fenológicos, lo cual está bajo el control de la disponibilidad de carbono y energía y de su distribución (Wolstenholme y Whiley, 1989) en respuesta a las condiciones medioambientales (Scholefield et al., 1985). Las hojas requieren alrededor de 40 días desde el desborre hasta la transición de sumidero a fuente (Whiley, 1990). Durante este período pueden competir por fotoasimilados con los frutos en desarrollo (Cutting y Bower, 1990).
El aguacate a lo largo del año puede tener uno o más ciclos vegetativos seguidos de un periodo de crecimiento radicular. Las raíces comienzan su crecimiento cuando el primer crecimiento vegetativo comienza a declinar. Posteriormente, comienza un segundo periodo de crecimiento vegetativo, restableciéndose de esta manera el equilibrio entre una fase de crecimiento radicular y otra vegetativa (Calabrese, 1992).

El aguacate es un frutal con un marcado efecto de alternancia de cosechas, donde condiciones climáticas óptimas para el cuajado producirán un año de elevada cosecha, que será seguido por un año de baja producción y, por el contrario, un año de condiciones climáticas que producen una elevada abscisión de flores y/o frutos, dará origen a un año de baja producción para, posteriormente, al año siguiente, aumentar la floración y con ello la cosecha (Lovatt, 2004).
La floración y el crecimiento de frutos utilizan grandes cantidades de carbohidratos, de modo que un déficit de éstos limitaría estos procesos y daría lugar a un año de escasez productiva. Dicho de otro modo, una alta carga frutal sería la principal causante de la reducción en la acumulación de carbohidratos durante el otoño y el invierno, dando por resultado una baja floración y un gran desarrollo vegetativo al año siguiente. Por el contrario, durante en un año de baja producción, los árboles acumularían niveles más altos de carbohidratos durante el otoño e invierno, dando como resultado una intensa floración (Paz Vega, 1997).

Scholefield et al. (1985) afirman que en otoño la concentración de carbohidratos es mínima, al mismo tiempo que tiene lugar una disminución de la actividad vegetativa, lo que se traduce en una menor competencia entre desarrollo vegetativo y reproductivo. Estos autores señalan que, de acuerdo con ello, la iniciación floral en el aguacate tiene lugar cuando los niveles de carbohidratos son mínimos, existiendo entonces una estrecha relación entre los niveles de carbohidratos y la iniciación floral. Los bajos niveles de carbohidratos podrían ser la causa del detenimiento del crecimiento vegetativo y ser este factor el determinante, en mayor grado, de la iniciación floral.
En el aguacate, los crecimientos vegetativos ocurren justo cuando los niveles de reservas en las ramas son muy bajos. Los niveles más altos de reservas de almidón en la madera se encuentran en invierno, coincidiendo con la detención del crecimiento vegetativo. Estas reservas descienden rápidamente durante la floración, para encontrarse en sus niveles más bajos durante el verano, momento en el que ocurre la abscisión de frutos (Whiley y Wolstenholme, 1990). Esta acumulación de almidón en invierno determina el mantenimiento del fruto en el árbol.
Scholefield et al. (1985) afirman que en otoño la concentración de carbohidratos es mínima, al mismo tiempo que tiene lugar una disminución de la actividad vegetativa, lo que se traduce en una menor competencia entre desarrollo vegetativo y reproductivo. Estos autores señalan que, de acuerdo con ello, la iniciación floral en el aguacate tiene lugar cuando los niveles de carbohidratos son mínimos, existiendo entonces una estrecha relación entre los niveles de carbohidratos y la iniciación floral. Los bajos niveles de carbohidratos podrían ser la causa del detenimiento del crecimiento vegetativo y ser este factor el determinante, en mayor grado, de la iniciación floral.
En el aguacate, los crecimientos vegetativos ocurren justo cuando los niveles de reservas en las ramas son muy bajos. Los niveles más altos de reservas de almidón en la madera se encuentran en invierno, coincidiendo con la detención del crecimiento vegetativo. Estas reservas descienden rápidamente durante la floración, para encontrarse en sus niveles más bajos durante el verano, momento en el que ocurre la abscisión de frutos (Whiley y Wolstenholme, 1990). Esta acumulación de almidón en invierno determina el mantenimiento del fruto en el árbol.
En las inflorescencias indeterminadas, las hojas compiten por fotoasimilados con las flores y el fruto en desarrollo hasta que alcanzan 2/3 de su expansión total (Whiley, 1990).Una vez que los frutos comienzan a desarrollarse, la dirección del transporte de fotoasimilados cambia en favor del crecimiento del fruto (Ho, 1992).

Un hecho adicional que se utiliza para apoyar la hipótesis de que la disponibilidad de carbohidratos podría ser el regulador en el ciclo de la alternancia productiva es el gran contenido en aceites del aguacate, cuya acumulación es energéticamente muy costosa. Si lo comparamos con otros frutos, tales como manzanas, naranjas y plátanos, las cantidades de azúcares que almacenan son mucho menores, pero su alto contenido en aceite podría explicar la drástica reducción de las reservas de carbohidratos en un año de alta producción.
También se acepta que la alternancia productiva está relacionada con el papel de los reguladores de crecimiento (PGRs) producidos por la semilla en la inhibición de la floración del año siguiente (Paz Vega, 1997). En el aguacate, el ácido giberélico producido por el endocarpio del fruto se ha relacionado con el descenso de la floración y la producción de fruta (Martens et al., 1994). El papel de las reservas de carbohidratos queda, bajo este punto de vista, en entredicho (Liu et al., 1999).
Scholefield et al. (1985) mencionan que el cese del crecimiento vegetativo se relaciona con una disminución en la producción de giberelinas, permitiendo la iniciación de yemas florales, y, por otra parte, los frutos se caracterizan por su intensa demanda metabólica.
En general, podemos concluir que muchos factores hacen difícil el estudio de la alternancia de cosecha y su relación con el crecimiento vegetativo, por ejemplo, factores nutricionales, hormonales y climáticos y otros factores derivados de reacciones endógenos implicadas en la competencia entre órganos en crecimiento. Lo que si podemos es prevenir que su efecto se acentúe actuando en aquellos factores que sí podemos controlar, como por ejemplo los nutricionales.
Por último, cabe destacar que el almidón y los azúcares solubles son los carbohidratos de reservas predominantes para disponer de energía para el crecimiento (Dey y Dixon, 1985).
RECOMENDACIÓN
Por todo lo expuesto, desde el departamento técnico de Cultifort, se recomienda trabajar nutricionalmente durante las etapas de mayor demanda del aguacate, de manera que el crecimiento vegetativo y la floración y cuajado de frutos, se vean tan solo afectados por aquellos factores que escapan al control humano.

Para más información sobre cultivo del aguacate: consejo de un experto otro artículo que os pueden interesar tratamientos preventivos frente a la caída de fruto
Desde Cultifort se ofrece una amplia gama de bioestimulantes, correctores de carencias, enmiendas orgánicas y otros productos especiales, para satisfacer la demanda del aguacate en cada uno de sus estados fenológicos.
En la fase fenológica actual, crecimiento de brotes y frutos, hay que tener en cuenta lo expuesto anteriormente, que existe una alta competencia por fotoasimilados o carbohidratos entre los frutos en crecimiento y las hojas y brotes jóvenes. Esta competencia, además, puede provocar un incremento de la caída fisiológica de frutos si no se controla adecuadamente. Es, por tanto, muy recomendable corregir cualquier tipo de carencia nutricional y trabajar a nivel de bioestimulación en esta etapa tan crítica.
Como base para este periodo, se recomienda mejorar las condiciones edáficas, pues ya hemos visto que para que el crecimiento vegetativo sea eficiente, éste tiene que ir ligado a un buen desarrollo radicular. MICROVITAL – L, formulación líquida rica en ácidos orgánicos, moléculas flavonoides, magnesio y micronutrientes, es un activador biológico del suelo de origen vegetal que además de mejorar los parámetros físico-químicos, actúa como producto prebiótico, estimulando el crecimiento y desarrollo de la microfauna edáfica. Estos efectos se consiguen gracias a:

- Los ácidos orgánicos, en formas directamente asimilables por la planta (a diferencia de otros ácidos orgánicos)
- Y moléculas flavonoides, que como polifenoles que son, poseen propiedades antioxidantes, ayudando a la planta a permanecer fotosintéticamente más activa, retrasando la senescencia foliar y ejerciendo una acción protectora frente a distintos tipos de estrés abiótico, como puede ser por ejemplo la radiación ultravioleta. Además poseen propiedades de quelación de hierro y otros metales, por lo que el uso de MICROVITAL – L contribuye a la movilización de nutrientes presentes en el suelo.
Por si fuera poco, MICROVITAL – L, corrige y previene carencias de magnesio, boro, hierro, manganeso y zinc. Haciendo que la planta permanezca más verde, metabólicamente más activa, con un mejor desarrollo… en general, más sana y más fuerte.
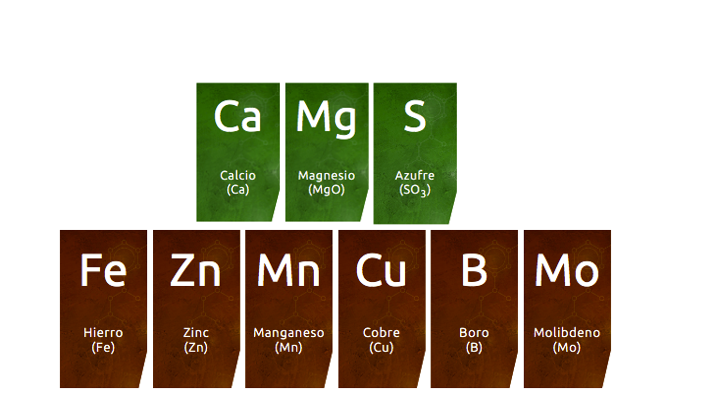
Dentro de la gama de correctores de carencias, Cultifort dispone de todo tipo de soluciones nutricionales: magnesio, calcio, hierro, boro, zinc, manganeso, molibdeno, etc., formulando todos ellos con la tecnología más eficiente para su correcta y máxima asimilación, utilizando distintos quelatos y agentes complejantes, junto con carbohidratos, poliácidos orgánicos, ácidos policarboxílicos, polisacáridos y azúcares reductores.
A nivel de bioestimulación, FOLITON es un bioestimulante con alto contenido en L-aminoácidos proteinogénicos, tanto libres como combinados en forma de péptidos y polipéptidos, que permiten ahorrar energía al aguacate en la formación de proteínas, especialmente cuando existen altos requerimientos nutricionales. Se trata de una formulación líquida de rápida asimilación y translocación en la planta. Su aplicación representa una fuerte estimulación del metabolismo vegetal en momentos en que se requiera aumentar la actividad vegetativa. La acción conjunta de los componentes de su formulación favorecen la síntesis de proteínas e hidratos de carbono, promueven el inicio de la actividad fisiológica de la planta y estimulan la formación de hojas y flores, reduciendo la relación competencial entre ellas.

SPIRALIS Long Life es una formulación de ácidos orgánicos especiales junto con un complejo de péptidos seleccionados, relacionados con microalgas rojas (Gellidium) y verdes (Spirulina). Su funcionalidad se basa en inducir y potenciar el incremento endógeno local y sistémico de moléculas con alta capacidad defensiva (alcaloides, tioninas, fitoalexinas, proteínas PR, etc.). Por otro lado, induce cambios estructurales en las paredes celulares de las plantas a nivel de su lignificación, constituyendo de este modo una barrera física frente al estrés abiótico. Estimula especialmente los niveles de respuesta de Resistencia Sistémica Adquirida (RSA) y Resistencia Sistémica Inducida (RSI) de la planta frente a hongos y bacterias, entre otros. Su uso se recomienda previo a situaciones de riesgo y/o mientras se mantengan las condiciones favorables para el desarrollo de enfermedades. Su efecto antifúngico y antibacteriano de amplio espectro, a nivel de prevención, y su efecto sobre la mejora de la vida post-cosecha de los frutos, convierten a SPIRALIS Long Life en uno de los productos más completos y técnicos del catálogo de Cultifort.
Pero también es conveniente proteger al aguacate frente a distintos tipos de estrés que puedan producirse durante esta fase, bien sea a nivel radicular como a nivel foliar.

OXIFORT es un producto que aplicado junto con el agua de riego, libera oxigeno lentamente, mejorando la aireación en la zona radicular, deteniendo los procesos de asfixia radicular y el desarrollo de microorganismos anaerobios. Además mejora la estructura del suelo, dándole esponjosidad, haciendo que el aire y el agua circulen mejor por el espacio poroso. Aumenta el rendimiento de la fertilización, especialmente la nitrogenada y favorece el desarrollo de microorganismos beneficiosos que participan en la transformación y movilización de nutrientes del suelo. OXIFORT es muy importante porque previene el estrés causado por condiciones de asfixia radicular. Recordemos que el cultivo del aguacate es extremadamente sensible al exceso de humedad en el suelo. Las condiciones de asfixia radicular se producen, bien por un mal manejo del riego o bien por un exceso de lluvia en una época de menor demanda hídrica. En cualquier caso, existe un periodo de 90 – 100 días aproximadamente, posteriores a floración y cuajado de frutos, en los que este cultivo es especialmente sensible al exceso de humedad, pudiendo sufrir un estrés por asfixia radicular que provoque una caída masiva de frutos de manera directa. Para evitar en la medida de los posible que se den tales situaciones, se recomienda realizar las plantaciones de aguacate en suelos sueltos con un volumen de macroporos superior al 17% (24% es lo ideal), de buen drenaje, con una profundidad mínima, planificando bien las salidas de agua de la parcela evitando acumulaciones en las partes más bajas y realizando un buen manejo del riego, optimizando las aportaciones de agua y la oxigenación del suelo.
CultisilK es un formulado a base de silicato potásico que además incorpora aminoácidos libres. Por un lado, el silicio va a crear una capa protectora sobre la planta para amortiguar el régimen térmico frente a un exceso de temperatura, protegiendo, además, frente a los golpes de sol; por otro lado, el potasio va a actuar como regulador del buen comportamiento estomático, ayudando a mantener la turgencia celular y evitando así la pérdida excesiva de agua por transpiración; y por último, los aminoácidos libres, van enfocados a reforzar la síntesis de clorofila y a activar la fotosíntesis, evitando la interrupción en la síntesis de asimilados y asegurando que no se acentúen la relaciones competenciales entre los frutos y los brotes en crecimiento. Como fuente de potasio y silicio, mejora el crecimiento de la planta y refuerza su resistencia a factores ambientales potenciando las autodefensas. El silicio es un elemento estructural que refuerza la pared celular, fortaleciendo la sustentación física del vegetal y protegiéndolo del ataque de agentes externos. Además, tiene sinergias con el calcio, el magnesio y el potasio, mejorando su absorción y transporte en la planta.
Resumiendo, en Cultifort ofrecemos una amplia gama de soluciones nutricionales con funciones específicas destinadas a mejorar los rendimientos del cultivo de una manera eficaz y ecológica. Además en nuestro catálogo sólo se incluyen productos “residuo cero”.
REFERENCIAS
Alcaraz, M.; Thorp, T. and Hormaza, J. (2013). Phenological growth stages of avocado (Persea americana) accordingto the bbch scale. Scientia Horticulturae, 164, 434-439. https://doi.org/10.1016/j. scienta.2013.09.051.
Calabrese, F. 1992. El aguacate. Madrid. Ediciones Mundi-Prensa. 249 p.
Coutanceau, M. 1964. Fruticultura. España, Ediciones de Occidente. 108p.
Cutting, J.G. and Bower, J.P. 1990. Spring vegetative flush removal: the effect on yield size, fruit mineral composition and quality. South African Avocado Growers´ Association Yearbook 13, 33-34.
Dey, P.M. and Dixon, R.A. 1985. Biochemistry of Storage Carbohydrates in Green Plants, 1−51. Dey, P.M. and Dixon, R.A. (Eds.). Academic Press, London.
Ho, L.C. 1988. Metabolism and compartmentation of imported sugars in sink organs in relation to sink strength. Ann. Rev. Plant Physiol. 39, pp. 355–378.
Ho, L. C. 1992. Fruit growth and sink strength. In: Fruit set production: Aspects of development, environmental physiology and ecology. Marshall, C., and Grace J. (Eds.). Cambridge Univ. Press. Great Britain. pp: 101-124.
Galán-Sauco, V. 1990. Los frutales tropicales en los subtrópicos (Aguacate-mango-LitchiLogan). España. Ediciones Mundi-Prensa.133p.
Liu, X., Robinson, P., Madore, M., Witney, G. and Arpaia, M. 1999a. Hass avocado carbohydrate fluctuations. I. Growth and phenology. Journal of the American Society for Horticultural Science 124: 671-675.
Lovatt, C. 2004. Eliminating alternate bearing of “Hass” Avocado. Proceeding of the California Avocado Research Symposium, October 30, 2004. University of California, Riverside. California Avocado Commission. Pages 89-95.
Martens, D.A., S. Luck, and W.T. Frankenberger, Jr. 1994. Role of plant growth regulators in vegetative spring flush, flowering and fruit drop in avocado (Persea americana Mill.). Spec. Rpt. Calif. Avocado Dev.Org., Calif. Avocado Soc., Saticoy.
Meyer,B. 1960. Introducción a la fisiología vegetal. Buenos Aires, Eudeba. 57Op.
Paz-Vega, S. 1997. Alternate bearing in the avocado (Persea americana mill). California Avocado Society Yearbook 81: 117-148.
Scholefield, P.B., Sedgley M. and Alexander, D.McE., 1985. Carbohydrate cycling in relation to shoot growth, floral initiation and development and yield in theavocado. Scientia Horticulturae 25, 99-110.
Thorp; T, and Sedgley, M. 1992. Shoot growth and tree architecture in range of avocado cultivars. In: Proceedings of Second World Avocado Congress. California 1992. pp. 237-240.
Whiley, A.W., 1990. CO2 assimilation of developing fruit shoots of cv Hass avocado (Persea americana Mili.) – A preliminary report. S.A. Avocado Growers’ Assn. Yrbk. 13,28-30.
Whiley, A.W. and Schaffer, B. 1994. Avocado. In: Schaffer, B., Anderson, P. (eds.). Handbook of Environmental Physiology of Fruit Crops, Vol 2. Subtropical and Tropical Crops. CRC Press, Boca Raton, Florida. Pp. 3-35.
Whiley A.W., Schaffer, B. and Lara, S.P. 1992. Carbon dioxide exchange of developing avocado (Persea americana Mill.) fruit. Tree Physiology 11, 85-94.
Whiley, A.W., and B.N, Wolstenholme. 1990. Carbohydrate managementin avocado trees for increased production. South African Avocado Growers Association Yearbook 13: 25 – 27.
Wolstenholme B. N. 1990. Resource allocation and vegetative reproductive competition: opportunities for manipulation in evergreen fruit trees. Acta Horticulturae 275:451-459.
Wolstenholme, B.N. and Whiley, A. W. 1989. Ciclos de carbohidratos y fenológicos como herramientas de manejo para los huertos de paltos. Asociación de Cultivadores de Paltos de Sudáfrica. Yrbk. 12: 33-37.